A post appeared on an I.D. Facebook group with this introduction:
Given the fact that he is a staunch Darwinist who fully accepts the basic tenets of the mainstream theory of evolution, you might expect Larry Moran (who, after all, did coin the derisive term “IDiots” in reference to Intelligent Design proponents) to have nothing but sneering contempt for Stephen Meyer.
Turns out, however, he met Stephen Meyer and the two got along surprisingly well, despite their deep ideological divide.
I went to the Moran’s blog and wanted to post his thoughts on Lawrence Krauss:
Krauss tried to hammer Meyer on the “ID is not science” issue using quotes from a judge based on things said by lawyers in the Dover trial….. Lawrence Krauss is an expert on cosmology but he’s very weak on biology. I know it’s common for physicists to think they are experts in everything but that’s just not true. It was demonstrated in last night’s debate.
In this public lecture at the University of California, Irvine, Professor Phillip E. Johnson explains how ambiguous terminology, faulty assumptions, and questionable rules of reasoning have transformed a theory which explains minor evolutionary change into a dogmatic naturalistic religion.
Major topics include:
Darwinism as a New Religion
Assumptions and Terminology
Darwinism and the Blind Watchmaker Hypothesis
Logical and Empirical Problems in Darwinian Theory
Rules of Reasoning in Modern Science
Some consider this to be one of Professor Johnson’s finest lectures based on his first book “Darwin on Trial.”
Are we One Nation under God or One Nation under Darwin? Phillip Johnson says this is the most important public debate to face our nation in the past 150 years. Either we are under God or we are not under God. Either God is real or God is imaginary. What we decide we are under as a nation will determine our moral authority for the future of our people and our country.
The inspiration for this lecture was the Pledge of Allegiance Case heard by the U.S. Supreme Court in 2004. In this case an atheist father was requesting that the words “under God” be removed from the Pledge of Allegiance his daughter was required to recite at her public school. There were several interesting twists to the case including the fact that he was an absent father who never married the mother, and both the mother and daughter were Christians and did not object to the “under God” phrase in the pledge. The father was a medical doctor, had also attended law school, and did an admirable job in arguing his own case before the California Court of Appeals for the 9th Circuit that ruled in his favor.
After the California court decision there was a public outcry from citizens across the nation and all 100 Senators in Congress registered disapproval of the decision. And yet Phillip Johnson argues, this decision was a logical progression of court rulings since the 1962 decision by the U.S. Supreme Court to prohibit prayer in our public schools. It was just one more step in the intentional secularization of America by our courts by removing any form of recognized religion from the public square, and leaving in its place only the hidden secular religion of naturalism.
Likewise the institution of marriage is under attack by the courts and the legal profession is already laying the ground work to redefine marriage from the union of a man and a woman, to the union of two people of the same sex, or a group of people, or possibly even between people and animals.
Ideas have consequences and the moral authority that “we the people” decide to put ourselves under in the future will have far reaching consequences for our nation.
What is the best way to discuss these issues in a pluralistic society? Phillip Johnson advocates the “Teach the Controversy” approach which has been endorsed by Congress in the Santorum language that was part of the “No Child Left Behind” Education Act of 2002.
The U.S. Senate voted 91-8 in favor of the Santorum Amendment to the “No Child Left Behind” Act. While the House of Representatives version of the bill did not include the amendment, a joint House-Senate committee included the following language from the original Santorum Amendment in its explanatory Conference Report: “The conferees recognize that a quality science education should prepare students to distinguish the data and testable theories of science from religious or philosophical claims that are made in the name of science. Where topics are taught that may generate controversy (such as biological evolution),
The below is a response to a conversation elsewhere on the WWW.
ARCHAEOPTERYX
I have a digital edition of Science Magazine and they allow me to read all the past articles (like this one, Archaeopteryx: Early Bird Catches a Can of Worms). In the article a wild eyed creationist… er… I mean, a respected evolutionist discusses how Archaeopteryx is more bird like, not the missing link between dino and bird.
In the article we find this:
feathers are 100% bird feathers;
hollow bones like birds;
It’s claws were perching claws (similar to the Bowerbird)…
doubts connected with dico/bird progression
…even the father of the modern “bird/dino” theory, John Ostrom, says this of recent revelations about Archaeopteryx:
“I’m just having a ball,” he said with a chuckle. “It sounds to me as if Alan [Feduccia] has presented a very good argument; I’m not sure he’s absolutely right, but I’m sure he’s on solid ground.”
Since the writing of the linked article at Science, more evidence (I will repeat, e-v-i-d-e-n-c-e) has come to light supporting the articles authored (Alan Feduccia) and curator of birds at the Smithsonian Institute, Storrs Olson:
bumps in the bone where feathers were connected (just like birds);
the avian lung was present in Archaeopteryx [pneumatized vertebrae and pelvis];
Cat Scans of the skull shows that the brain was birdlike, not dino-like (“Axial and appendicular pneumaticity in Archaeopteryx,” Proceedings of the Royal Society of London, Series B. 267:2501–2505, 2000);
similar inner ear findings (“The avian nature of the brain and inner ear of Archaeopteryx,” Nature 430(7000):666–669, 5 August 2004; Witmer, L.M, “Inside the oldest bird brain, perspective,” same issue, pp. 619–620);
In 2011, “the Royal Society’s Biology Letters, the researchers wrote that Archaeopteryx’s assignment to a dinosaur group earlier this year ‘was acknowledged to be weakly supported’, They constructed new cladograms that pictured Archaeopteryx with birds, and not with any dinosaurs, with a caption that reads, ‘Archaeopteryx robustly reinstated as the most basal bird’.” (“Likelihood reinstates Archaeopteryx as a primitive bird,” Biology Letters. Published online before print October 26, 2011).
Since other feathered “birds” have been found around the same time or earlier than Archaeopteryx, causing Alan Feduccia to quip, “You can’t be older than your grandfather” (Creation.com)… Nature has published an article pointing out that Archaeopteryx is JUST LIKE modern flightless birds.
And so it could have been losing its ability for flight (like modern birds have).
“We know Archaeopteryx was living on an archipelago during the Jurassic. And with its feathers and bones looking so much like modern flightless island birds, it just makes me wonder,” says…. Michael Habib, a biologist at the University of Southern California….
[….]
“Just because Archaeopteryx was the first feathered dinosaur found, doesn’t mean it has to play a central role in the actual history of the origins of birds,” says palaeontologist Thomas Holtz of the University of Maryland in College Park. “We have to remember it appears 10 million years or so after the oldest known bird-like dinosaurs and so our famous ‘first bird’ may really be a secondarily flightless one.”…
Recent events cast even further doubt on Archaeopteryx as a transitional form. If the claims of Sankar Chatterjee prove to be valid, then certainly Archaeopteryx could not be the ancestral bird, and dinosaurs could not be ancestral to birds. Chatterjee and his co-workers at Texas Tech University claim to have found two crow-sized fossils of a bird near Post, Texas, in rocks supposedly 225 million years old—thus allegedly 75 million years older than Archaeopteryx and as old as the first dinosaurs. Totally contrary to what evolutionists would expect for such a fossil bird, however, Chatterjee claims that his bird is even more bird-like than Archaeopteryx! In contrast to Archaeopteryx, this bird had a keel-like breastbone and hollow bones. In most other respects, it was similar to Archaeopteryx.14 If evolutionary assumptions are correct, this bird should have been much more reptile-like than Archaeopteryx. In fact, he shouldn’t even exist!
14.S. Weisburd, Science News, August 16, 1986, p. 103; Tim Beardsley, Nature 322:677 (1986).
But even if its classification waffles again, it is disqualified as an evolutionary ancestor for birds by the fact that scientists found a crow-size bird and extinct four-winged birds in rock layers designated to be below those containing Archaeopteryx.3,4
There is just as much [at best] evidence for this proposition as the next. “Devolution” — a loss of specificity, may be a more reasonable position to take via observed evidence. We see this all the time (directly below is an example from Lee Spetner’s new book), and EVOLUTION NEWS says that “looks like Archaeopteryx may have to be reclassified as a different sort of icon — symbolizing evolution by loss of function.” Oops.
Antibiotic Resistance
The evolution of antibiotic resistance has been for some time the Darwinists’ favorite example for “demonstrating” evolution (Common Descent). Superficially their case looks good. Antibiotics date only from about 1930 with the discovery of penicillin (Fleming 1929), followed by the development of a method to produce it with high yield (Chain et al. 1940). Antibiotics were first introduced to the public in 1942 to cure bacterial infection (Levy 1992, 4), and by the mid 1940s the first strains appeared of Staphylococcus resistant to penicillin (Fisher 1994, 15). Just a few years after antibiotics were introduced, resistant strains of the pathogens were found to have already evolved. As each new antibiotic was discovered and put into use against pathogenic bacteria, resistant strains soon followed. The argument then goes, with a wave of the hand, like this: If a small but significant evolutionary change like antibiotic resistance can evolve in only a few years, then surely in a million years huge evolutionary changes must occur. Darwinists expect this argument to support Common Descent.
An examination of the phenomenon of antibiotic resistance, however, shows it lends no support at all to Common Descent (Spetner 1997, 138143). Antibiotics are natural molecules produced by some microorganisms for the purpose of killing other hostile microorganisms. A microorganism that makes an antibiotic must, itself, be resistant to the antibiotic it makes. For this purpose it is typically endowed with a battery of genes that code for a resistance mechanism. Most useful antibiotics have come from soil bacteria (D’Costa et al. 2006). How bacteria have acquired this resistance initially is not known, nor can neo-Darwinian theory shed any light on it. Antibiotic resistance genes have been found to predate the use of antibiotics by at least many thousands of years (D’Costa et al. 2011). Moreover, bacteria are known to be able to transfer genetic material to other bacteria through HGT (see above). On occasion, copies of the genes for resistance can find their way from a type of bacterium that is normally resistant to a type that is not normally resistant. When that happens, the recipient bacterium becomes resistant. This is indeed evolution, but it is a limited evolution of the population-change type. It is not the Common-Descent type of evolution.
The resistance genes already exist in the biosphere. No new information has appeared in the biosphere through this type of evolution of antibiotic resistance. Common-Descent evolution cannot be achieved by this procedure even if it were repeated innumerable times in succession, because no new information would be built up. This method of evolving antibiotic resistance therefore lends no support for Common Descent.
Sometimes, however, antibiotic resistance can indeed appear through a random mutation — a DNA copying error, which would bring something new to the biosphere. This kind of change looks like it might satisfy the requirements for Common Descent, so I shall give a brief description of it here, although I have already dealt with it in my previous book.
As an example, let us look at how a bacterium acquires resistance to streptomycin through a random mutation. All cells, whether of bacteria or of plants or animals, contain organelles called ribosomes, whose function it is to make protein according to instructions from the DNA of a gene. Proteins are large molecules, consisting of long chains of small molecules called amino acids, and are essential to all living things. They function as enzymes, which catalyze all the chemical reactions in a cell — each chemical reaction catalyzed by a specific enzyme. Proteins can also serve as structural elements. Often, and maybe even always, a structural protein functions also as an enzyme. For an enzyme to perform its function, it must have a specific sequence of amino acids.
A ribosome is an organelle within a cell that manufactures protein. It makes a protein by putting together a chain of amino acids according to the instructions in the DNA. A segment of the DNA is transcribed into an RNA molecule that matches the DNA nucleotide by nucleotide. This RNA is called messenger RNA because it carries the DNA message to the ribosome. The ribosome translates the message in the DNA into amino acids according to the genetic code. Three nucleotides translate into one amino acid. Accordingly, the ribosome constructs a chain of amino acids to form a protein.
The antibiotic streptomycin, for example, acts on a bacterial cell by attaching to a ribosome at a site to which it matches, the way a key fits into a lock. When the streptomycin molecule attaches to this site, it interferes with the ribosome function and causes it to make mistakes leading to incorrect, dysfunctional or nonfunctional, protein. The errors it causes prevent the cell from growing, reproducing, and eventually from living. The important feature of streptomycin, and indeed of all other antibiotics, is that it kills bacteria but does not harm the mammalian host. Streptomycin kills the bacterial cells that are infecting you without killing your own cells. It discriminates between the cells of the bacteria and the cells of the host by its specific attachment to a matching site on the bacterial ribosome, a site not found on the host’s ribosomes.
A bacterium will gain resistance to streptomycin if a point mutation occurs in the gene coding for the protein in the ribosome, ruining the matching site, destroying the specificity of the protein, and preventing a streptomycin molecule from attaching. If the streptomycin cannot attach to the matching site, the bacterium is resistant. Just one mutation in the portion of the DNA coding for the matching site can mess up the site so the streptomycin cannot attach. It turns out that any one of several mutations in that portion of the DNA will grant the bacterium resistance (Gartner and Orias 1966). Note that this type of resistance is caused by a single random point mutation, but it cannot serve as an example of mutations that can support Common Descent. One cannot expect mutations destroying specificity, no matter how many of them there are, to build information and lead to Common Descent. Destruction of specificity does not add information — it destroys it. One cannot add information by destroying it, no matter how many times one repeats the process. I have previously (Spetner 1997) compared trying to build up information in this manner to the merchant who was losing a little money on each sale but thought he could make it up on volume. The acquisition of antibiotic resistance is indeed evolution, but only a limited form of it. It cannot lead to Common Descent.
No example of antibiotic resistance in bacteria adds information to the biosphere. To become resistant, the bacteria either pick up ready-made resistance genes from other bacteria or they undergo a mutation that destroys information. Antibiotic resistance cannot therefore be an evolutionary example that could support Common Descent because a chain of such mutations, no matter how long, does not add information and thus cannot lead to Common Descent. The Darwinists’ favorite example of evolution fails to pass muster.
End Notes
Chain, E. et al. (1940) Penicillin as a Chemotherapeutic Agent. Lancet 239: 226-228.
D’Costa, Vanessa M., Katherine M. McGrann, Donald W. Hughes, and Gerard D. Wright. (2006) Sampling the antibiotic resistome. Science 311: 374-377.
D’Costa, Vanessa M. et. al. (2011) Antibiotic Resistance is Ancient. Nature 477:457-461.
Fisher, Jeffrey A. (1994) The Plague Makers: How we are creating catastrophic new epidemics — and what we must do to avert them. New York: Simon & Schuster.
Fleming, A. (1929) On the antibacterial action of cultures of a Penicillium, with special reference to their use in the isolation of B. influenzae. British Journal of Experimental Pathology 10: 226-238.
Gartner, T. K. and E. Orias, (1966) Effects of mutations to streptomycin resistance on the rate of translation of mutant genetic information. Journal of Bacteriology 91: 1021-1028.
Levy, Stuart B. (1992) The Antibiotic paradox: How Miracle Drugs are Destroying the Miracle. New York: Plenum Press.
Spetner. L. M. (1997) Not by chance! Shattering the Modern Theory of Evolution. Brooklyn: Judaica Press.
A friend comments about the newer position on his Creation/Evolution Headlines saying one “paleontologist remarked, ‘We really need an improved understanding of how anatomy relates to these diverse behaviours, so we can better interpret the fossil record’.” Continuing he adds his thinking to the matter:
No one called Archaeopteryx a “feathered dinosaur” back then, because the phrase only came into vogue with the Chinese fossil discoveries. From Darwin’s day till recently, it was argued to be a transitional form between reptiles and birds. Evolutionists emphasized the reptilian traits (teeth, claws on the wings), and creationists emphasized the flight feathers and anatomy that seemed to show it capable of powered flight. They also pointed out that some living birds, like the hoatzin, have claws on their wings as juveniles. People saw what their biases wanted to see. Astronomer Fred Hoyle tried to prove it was a forgery. Today’s evolutionists use the “feathered dinosaur” label, but there is no guarantee that today’s consensus will not shift again. The new proposal it was secondarily flightless implies a win for creationists – it devolved from a fully-functional flying bird, just like some living birds with stunted wings have on the Galapagos Islands. Loss of function is not what Darwin needs!
Let’s think about Nature’s comment that the suggestion Archaeopteryx was losing the ability to fly “might have been considered madness” back in 1861 (actually, all the way from 1861 to just a few years ago). This tells us that if evolutionists consider something madness now, it might be considered sanity later. It further means that the sane ones could be the skeptics of the consensus, and the mad ones in the majority. Don’t be deterred, therefore, if you feel you have good evidence and arguments for your position when it runs counter to the consensus. It’s entirely possible for the intellectual majority to be suffering from delusions. “We really need an improved understanding … so we can better interpret the fossil record” – good advice, but it implies that understanding is lacking and interpretation is flawed. If they haven’t gotten it down after 152 years, don’t expect major improvements any time soon. They might just be secondarily clueless.
Here is a quote, and really, a definition of the general theory of evolution (GTE) that G.A. Kerkut defines in his older text, Implication of Evolution (second quote). Here Spetner calls it the neo-Darwinian theory (NDT), it more common name today. Here is Spetner’s relevant quote:
Neo-Darwinian Theory (NDT) is counterintuitive, and is acknowledged as such even by its supporters. All present-day life is assumed to have evolved from some primitive cell, and that cell was supposed to have formed itself from simple chemicals. Nobody seems to know how that cell came to be, but almost all biologists think they understand fairly well how evolution proceeded from that cell to all the life we see today.
There appears to be a vast amount of information contained in trees, fish, elephants, and people. Where did this information come from? It is said to have come from random mutations and natural selection. How can that work? Natural selection is supposed to be the magic that makes evolution happen, but all natural selection does is eliminate the less adaptive organisms and allow the more adaptive ones to survive and proliferate. Where do those more adaptive ones come from? Apparently, that’s what random mutations are supposed to accomplish.
So the information buildup required by Common Descent can come only from random mutations. That means that the buildup of information is a matter of chance. At each step of the evolutionary process, a mutation has to have occurred that grants the organism an advantage. The big question is: Is that reasonable? To see if it is, some people (including me) have made calculations of the probability of mutations building information.
We really don’t have all the data we need to make this calculation. But even if we make some conservative assumptions and give the benefit of all doubts to the Darwinian side, such calculations demonstrate that Common Descent is not reasonable. The Darwinists, however, do not accept these calculations as conclusive — they suggest alternative scenarios that might make the probabilities larger.
In his book Darwin’s Black Box, Michael Behe addressed the unreasonableness of Darwinian evolution. He described some biological systems as what he called “irreducibly complex.” By that he meant that these systems are composed of several critical components in such a way that the system cannot work unless all those components are in place. He then argued that the system could not evolve one small part at a time, because natural selection could not work on less than the whole system. Here, too, the Darwinians countered by suggesting scenarios in which natural selection might work, but again, the Darwinian scenarios are purely hypothetical.
Because the Darwinians can invent scenarios to address any challenge to their theory, they are not convinced by attempts to show that neo-Darwinian evolution cannot work. Therefore, I have concluded that it would be more productive to challenge them to show that it could work — challenge them to do more than just offer vague scenarios of how their theory might work, but to show by calculation that the probability of it working is reasonably high. This is a challenge they must meet to establish their theory on a scientific basis. They have never met this challenge and they cannot. They cannot show that the events they claim to have produced Common Descent have a high enough probability to justify their claim. Their inability to establish the theory of Common Descent means that Common Descent is not an established theory. This is one of the main points of this book.
I cannot overemphasize the importance of probability calculations. NDT is not like Newton’s theory of mechanics, whose equations describe the motion of a physical body under a force. Nor is it like Maxwell’s theory of electromagnetism, whose equations describe the effects of electric and magnetic fields on electric charges. These theories are checked against experiment by solving those equations. NDT describes evolution as the result of random mutations that may or may not yield an adaptive phenotype. These are chance events. The theory can be checked only by calculating the probabilities of the required events to see if they are reasonably large. The theory has not been shown to have passed this test and is therefore not a valid theory. Whatever evidence is given for Common Descent is circumstantial. Circumstantial evidence cannot stand alone. It needs to have a theory tying the evidence to the conclusion. But instead of a theory, imaginary scenarios are offered to suggest how evolution might work. No calculations of probabilities are made.
[….]
Common Descent is a key component of an agenda advocating a natural origin of life. The effort to demonstrate the possibility of such a natural origin is usually divided into two parts: (1) abiogenesis, the origin of a simple life form from naturally occurring chemicals, and (2) the evolution of all life from that single simple beginning. It turns out, however, there is no good evidence for either of these two parts.
This is an extended quote from a MUST HAVE resource, The Genesis Account ~ for the seminarian or creation enthusiast (maybe a graduated-seminary student continuing his or her studies? of which I am). Again, this is not an introduction to the topic of “creation,” but rather a serious in-depth review of history, science, language, culture, theology and the Scriptures. [Two videos added to deepen understanding of molecular machines.]:
DNA: INFORMATION STORAGE, RETRIEVAL, AND TRANSMISSION
Not only do living things constitute enormously complex machines, they also contain the ‘instruction manual’ to build them—a sort of ‘recipe book’ programmed on DNA, the famous ‘double helix’ molecule (deoxyribonucleic acid). The information is stored in four different chemical ‘letters’: A T, C, and G.33 Plants and microbes also use the same DNA coding system.
These ‘letters’, or bases, have remarkable chemical properties. They are flat molecules, which allows them to stack as ‘rungs’ on a spiral staircase. This provides some stability, although see below. The letters of DNA have another vital property due to their structure, which allows information to be transmitted: A pairs only with T, and C only with G, due to the chemical structures of the bases—the pair is like a rung or step on a spiral staircase. This means that the two strands of the double helix can be separated, and new strands can be formed that copy the information exactly. The new strand carries the same information as the old one, but instead of being like a photocopy, it is in a sense like a photographic negative.
The copying is far more precise than random or laboratory chemistry could manage, because there is editing (proof-reading and error-checking) machinery, again encoded in the DNA. This machinery keeps the error rate down to less than one error per l07-108 letters).34, But how would the information for editing machinery be transmitted accurately before the machinery was in place? Lest it be argued that the accuracy could be achieved stepwise through selection, note that a high degree of accuracy is needed to prevent ‘error catastrophe’—the accumulation of ‘noise’ in the form of junk proteins specified by the damaged DNA.
DNA is unstable
DNA is a very complicated molecule, and actually a very unstable one. DNA researchers often need to store it in liquid nitrogen, at about —200°C, and even that frigid temperature doesn’t entirely stop breakdown. A recent paper on DNA stability estimates that, even when preserved in bone, it would be completely disintegrated in 22,000 years at 25°C, 131,000 years at 15°C, 882,000 years at 5°C; and 6.83 million years at —5°C.35 One article reported:
There is a general belief that DNA is ‘rock solid’—extremely stable,” says Brandt Eichman, associate professor of biological sciences at Vanderbilt, who directed the project. “Actually DNA is highly reactive. On a good day about one million bases in the DNA in a human cell are damaged.36
Fortunately, in our cells, we have many elaborate repair machines to undo this chemical damage.37 But most skeptics believe that life evolved in a primordial soup,38 which would have lacked such machines (not to mention the lack of any evidence that it existed at all39). So even if DNA managed to form spontaneously somehow, it would not have survived long.40
Encyclopedic information store
But even more important than the chemistry is the enormous information content of the ‘recipe’ stored on this DNA. This is transmitted from one generation to the next, so that living things reproduce ‘after their kinds’ (cf. Genesis 1). Leading atheist Richard Dawkins himself admits:
[T]here is enough information capacity in a single human cell to store the Encyclopedia Britannica, all 30 volumes of it, three or four times over.41
Nowadays we would say that each of our cells—and there are about a hundred trillion in the human body—contains about three gigabytes of information. Even the simplest living creature, the tiny germ Mycoplasma, has about 600 kilobytes.42 And even this seems incredibly highly compressed. Some bioengineers, led by Stanford University’s Markus Covert, succeeded in modelling this ‘simple’ germ with computers.43 One report on trying to model the processes involved in one cell division for this cell stated:
What’s fascinating is how much horsepower they needed to partially simulate this simple organism. It took a cluster of 128 computers running for 9 to 10 hours to actually generate the data on the 25 categories of molecules that are involved in the cell’s lifecycle processes.44
To return to the Britannica, the information is in the form of ink molecules on paper. But nothing in the ink molecules themselves made them form into the letters, words, phrases, sentences, and paragraphs of the Encyclopedia. It certainly wasn’t produced by an ink spill. Rather, the information was imposed on the ink by an outside intelligent source (or a program ultimately programmed by an intelligent mind).
But here is the connection with living things. There is likewise nothing in the chemistry of DNA’s letters themselves that would make them join up in predetermined ways, any more than forces between ink molecules make them join up into letters and words. Michael Polanyi (Hungarian: Polányi Mihaly, 1891-1976), a former chairman of physical chemistry at the University of Manchester (UK) who turned to philosophy, confirmed this:
As the arrangement of a printed page is extraneous to the chemistry of the printed page, so is the base sequence in a DNA molecule extraneous to the chemical forces at work in the DNA molecule. It is this physical indeterminacy of the sequence that produces the improbability of any particular sequence and thereby enables it to have a meaning—a meaning that has a mathematically determinate information content.45
Just as the Britannica had intelligent writers to produce its information, so it is scientific to believe that the information in the living world likewise had an original Writer.46
CELL MACHINES
Alex Williams, who was Australian representative to a research program coordinated by the UN’s International Atomic Energy Agency, explained this further in applying this to the cell machinery:
Polanyi pointed to the machine-like structures that exist in living organisms. … Just as the structure and function of these common machine components cannot be explained in terms of the metal they are made of, so the structure and function of the parallel components in life cannot be reduced to the properties of the carbon, hydrogen, oxygen, nitrogen, phosphorus, sulphur and trace elements that they are made of. There are endless examples of such irreducible structures in living systems, but they all work under a unifying principle called `autopoiesis’. Autopoiesis literally means ‘self-making’ (from the Greek auto for self, and the verb poieo meaning ‘I make’ or ‘I do’) and it refers to the unique ability of a living organism to continually repair and maintain itself—ultimately to the point of reproducing itself—using energy and raw materials from its environment.47
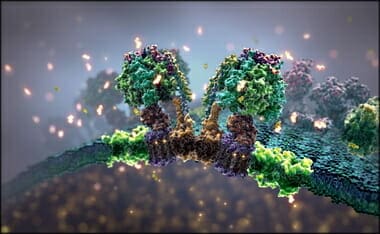
For example, for creatures to live at all, they need energy. This energy is supplied by a molecule called ATP.48 In fact, the human body generates—and consumes—about its own weight of ATP every day. Nowadays we know that ATP is produced by the world’s tiniest motor, ATP synthase.49[First Video Below] This is only 10 nm across by 8 nm high—so tiny that 1017 would fill the volume of a pinhead. Even the journal Nature called these motors “Real engines of creation”.50 Recent work shows that it’s also the most efficient motor in the world—in fact as efficient as the laws of physics allow-100%.51
But for ATP synthase to work as a builder of ATP, it must be in a membrane that provides an electric potential that drives a proton current to power it. Without this membrane, ATP synthase would operate in reverse, as an ATP destroyer (all enzymes are reversible).
There are many more nano-machines in living things, for example, the kinesin linear motor [Second Video Below] that ‘walks’ along miniature highways in the cell, called microtubules. In 8-nm steps (125,000 per millimetre), at a rate of 100 steps per second, it delivers protein packages to the right place in the cell. It knows where to go because of address labels on the packages! Kinesin is powered by ATP: one molecule per step.52
Above Video
(Creation.com) This animated sequence shows the ATP Synthase enzyme in operation. The animation is based on an incredible series of scientific discoveries. Only the colours show artistic licence.
ATP, or Adenosine Tri-Phosphate, is the energy currency of the cell. ATP is produced by a tiny molecular rotary motor, rotating at up to 7,000 rpm. These are so small that 100,000 would fit side-by-side in a millimetre. A current of protons drives the motor, unlike man-made electric motors, which use electrons.
This portion of the enzyme is where Adenosine Di-Phosphate is combined with a phosphate ion, in the presence of a catalyst to produce ATP which is then released, making way for the next cycle. A top view of the enzyme shows the sequential operation. Almost every bio-chemical process in your body requires ATP.
Such a nano-machine exhibits all the characteristics of super-intelligent design. ATP is vital for life and many of these motors were needed before the first living cell could exist. An evolutionary impossibility!
(Creation.com) Inside a living cell is an amazing transportation system. Proteins have to be delivered to the correct part of the cell to perform their intended functions. This animation, based on a lot of clever research over a number of years, shows how it happens.
Highways, made of microtubules, are assembled by interlocking proteins, each manufactured in accordance with the coded instructions on the cell’s DNA. Marching along a microtubule is the kinesin motor, the hero of our story, carrying a huge sack of proteins to be delivered to a pre-determined place in the cell. Here the proteins will be released to fulfil their functions.
A kinesin linear motor uses one ATP to provide the energy for each step and takes 125,000 steps to cover one millimetre!
This amazing machine shows all the hallmarks of design!
Information is meaningless unless we understand its language. The Britannica is little use unless we know English. For example, ‘gift’ in English means a present, but in German it means poison. The wrong convention can mean the wrong message. One German immigrant to New Zealand told me that in his first year, he thought that New Zealanders had macabre customs—at Christmas, they tried to poison their relatives and best friends!
The DNA code is also a language: three DNA ‘letters’ code for one protein `letter’. This language requires many different decoding machines to read it, including the ribosome.53 However, the instructions to build this decoding machinery are themselves stored on the DNA, thus producing a vicious circle, or chicken-and-egg problem.
Also, most of these processes use energy, supplied by ATP, produced by the nano-motor ATP synthase. But the ATP synthase motor can’t be produced without instructions in the DNA, read by decoding machinery using ATP… a three-way circle, or perhaps an egg-nymph-grasshopper problem.
Furthermore, DNA needs many machines for copying itself, but it also codes the instructions to build its own copying machines. But these instructions to build copying machines can’t be passed on without the copying machines already present … .
Multiple languages in DNA
In fact, there is more than one language involved. Recently, another code was discovered: the ‘splicing code’ that controls how different parts of the DNA are chopped out and spliced together; an editing process.54 This enables a single gene to encode multiple proteins, and explains why humans have only about 21,000 genes yet make up to a million proteins, which surprised those who decoded the human genome. For example, thanks to studies of the splicing code, researchers found that “three neurexin genes can generate over 3,000 genetic messages that help control the wiring of the brain,” according to co-discoverer Brendan Frey.55,56 This also involves a complex machine called a spliceosome. One paper was tellingly entitled, “Mechanical devices of the spliceosome: motors, clocks, springs, and things.”57
Multiple codes are an even bigger problem for evolution, as geneticist John Sanford (b. 1950), the inventor of the gene gun, pointed out:
Most DNA sequences are poly-functional and so must also be poly-constrained. This means that DNA sequences have meaning on several different levels (poly-functional) and each level of meaning limits possible future change (poly-constrained). For example, imagine a sentence which has a very specific message in its normal form but with an equally coherent message when read backwards. Now let’s suppose that it also has a third message when reading every other letter, and a fourth message when a simple encryption program is used to translate it. Such a message would be poly-functional and poly-constrained. We know that misspellings in a normal sentence will not normally improve the message, but at least this would be possible. However, a poly-constrained message is fascinating, in that it cannot be improved. It can only degenerate. Any misspellings which might possibly improve the normal sentence will be disruptive to the other levels of information. Any change at all will diminish total information with absolute certainty….
The poly-constrained nature of DNA serves as strong evidence that higher genomes cannot have evolved via mutation/selection except on a trivial level.58
The multiple languages are consistent with the almighty Creator God revealed in the Bible.
ATHEIST CONUNDRUM
The origin of the cell’s information, and its decoding machinery, is thus a huge problem for atheists. Astrophysicist and evolutionist Paul Davies (b. 1946) says:
We now know that the secret of life lies not with the chemical ingredients as such, but with the logical structure and organisational arrangement of the molecules. … Like a supercomputer, life is an information processing system. … It is the software of the living cell that is the real mystery, not the hardware.59
But he does nothing to solve this mystery. Instead he asks, “How did stupid atoms spontaneously write their own software?”, and answers, “Nobody knows …” and admits: “There is no known law of physics able to create information from nothing.”
However, this makes good sense if the information came from a Master Programmer, as revealed in the Bible.
Foot Notes
Adenine, cytosine, guanine and thymine. They are part of building blocks called nucleotides, which comprise the sugar deoxyribose, a phosphate and a base. In RNA, uracil (U) substitutes for thymine and ribose substitutes for deoxyribose.
Kunkel, T.A., DNA Replication Fidelity, J. Biological Chemistry 279:16895-16898, 23 April 2004
Allentoft, M.E. et aL, The half-life of DNA in bone: measuring decay kinetics in 158 dated fossils, Royal Society B 279(1748):4724-4733,7 December 2012.
Salisbury, D.F., Newly discovered DNA repair mechanism, Science News, com, 5 October 2010.
For problems with materialistic ideas that life evolved from non-living chemicals, see com/origin and Sarfati, J., By Design, ch. 11, 2008.
Brooks, J., and Shaw, G. point out, “If there ever was a primitive soup, then we would expect to find at least somewhere on this planet either massive sediments containing enormous amounts of the various nitrogenous organic compounds, acids, purines, pyrimidines, and the like; or in much metamorphosed sediments we should find vast amounts of nitrogenous cokes. In fact no such materials have been found anywhere on earth.” Origins and Development of Living Systems, 359, 1973.
Many skeptics believe that life started with a similar molecule called RNA (ribonucleic acid). But this is even less stable than DNA, and so are its building blocks such as the sugar ribose. John Horgan admits in ‘Scientists don’t have a clue how life began’ above, “But the ‘RNA-world’ hypothesis remains problematic. RNA and its components are difficult to synthesize under the best of circumstances, in a laboratory, let alone under plausible prebiotic conditions…. The RNA world is so dissatisfying that some frustrated scientists are resorting to much more far out—literally—speculation.” For those interested in chemistry, more chemical problems with ‘RNA World’ ideas can be found at creation.com/rna.
Dawkins, C.R., The Blind Watchmaker: Why the evidence of evolution reveals a universe without design, 115, 1986.
Fraser, C.M., et al., The minimal gene complement of Mycoplasma genitalium, Science 270(5235):397-403, 1995; perspective by Goffeau, A., Life with 482 Genes, same issue, pp. 445446. They reported 582,000 DNA bases or ‘letters’. Other reports have a different number, but all within the same ball park. For simplicity, I am treating each DNA ‘letter’ as a ‘byte’ of information, which is ‘in the right ball park’.
Karr, J.R. et al., A whole-cell computational model predicts phenotype from genotype, Cell 150(2):389-410, 20 July 2012.
Madrigal, A.C., To model the simplest microbe in the world, you need 128 computers, com, 23 July 2012.
Polanyi, M., Life’s irreducible structure, Science 160:1308-1312, 1968.
See also Sarfati, J., DNA: marvellous messages or mostly mess? Creation 25(2):26-31, 2003.
J.. ATP: The perfect energy currency for she cell. CRSQ 36(1):2-10, 1999.
Updated from J. Sarfati, Design in living organisms (motors), Creation 12(1):3-5, 1998, written not long after the original discovery of the motor was published, and the Nobel Prize was awarded to the discoverers. A more up-to-date layman’s article is Thomas, B., ATP synthase: Majestic molecular machine made by a Mastermind, Creation 31(4):21-23, 2009; creation.com/atp-synthase links to an animation made by CMI.
Block, S., Real engines of creation, Nature 386(6622):217-219, 1997; Comment on Hiroyuki Noji et al., Direct observation of the rotation of F1-ATPase, same issue, pp. 299-302.
Shoichi Toyabea a , Thermodynamic efficiency and mechanochemical coupling of F1-ATPase, PNAS 13 October 2011.
A lucid lay explanation is Calvin Smith, Incredible kinesin! Biological ‘robots’ will blow your mind!, com/incredible-kinesin, 26 June 2012. This contains animations of both kinesin and ATP synthase motors.
Sarfati, J., com/message, 2003, links to animations of a number of DNA decoding machines, including RNA polymerase, the ribosome, the t-RNA ‘adaptor’ and the chaperonin folding machine; as well as the ATP synthase motor
Barash, Y. et al., Deciphering the splicing code, Nature 465:53-59, 2010.
Cantin, P., Researchers crack ‘Splicing Code’, solve a mystery underlying biological complexity, com, 5 May 2010.
Geneticist Dr Robert Carter explains this lucidly in, Splicing and dicing the human genome: Scientists begin to unravel the splicing code, com/splicing, 1 July 2010.
Staley, J.P. and Guthrie, C., Cell 92(3):315-326, 1998
Sanford, J.C., Genetic Entropy & the Mystery of the Genome, FMS Publications, 3rd Ed., p. 131-133, 2008.
Davies, P. Life force, New Scientist 163(2204):27-30, 18 September 1999.
On this episode of ID the Future Casey Luskin interviews Caroline Crocker, who shares about her experience during the filming of “Expelled: No Intelligence Allowed” and how she first became interested in the debate over origins. What was it like for Dr. Crocker when she was expelled for questioning Darwinian evolution (and having Ben Stein write the Foreword for her new book, Free to Think)? Listen in and find out. Buy her book: Free to Think: Why Scientific Integrity Matters
The Real Story of How Caroline Crocker Was Expelled from George Mason University. Are scientists free to think and follow the evidence wherever it leads? On this episode of ID the Future, Casey Luskin interviews Caroline Crocker, president of the American Institute for Technology and Science Education and author of Free to Think: Why Scientific Integrity Matters. Dr. Crocker was famously expelled from her job at George Mason University. Listen in as she shares stories about her inspiring student and reveals details in her case for the first time.
On this episode of ID the Future, Caroline Crocker is interviewed by Casey Luskin about the principles of academic freedom in education. Listen in as Dr. Crocker shares from her experience in the classroom at George Mason University and how all the problems she navigated there had a common thread: the lack of integrity in science.
Intelligent design (ID) is the view that it is possible to infer from empirical evidence that “certain features of the universe and of living things are best explained by an intelligent cause, not an undirected process such as natural selection” [1] Intelligent design cannot be inferred from complexity alone, since complex patterns often happen by chance. ID focuses on just those sorts of complex patterns that in human experience are produced by a mind that conceives and executes a plan. According to adherents, intelligent design can be detected in the natural laws and structure of the cosmos; it also can be detected in at least some features of living things.
Greater clarity on the topic may be gained from a discussion of what ID is not considered to be by its leading theorists. Intelligent design generally is not defined the same as creationism, with proponents maintaining that ID relies on scientific evidence rather than on Scripture or religious doctrines. ID makes no claims about biblical chronology, and technically a person does not have to believe in God to infer intelligent design in nature. As a theory, ID also does not specify the identity or nature of the designer, so it is not the same as natural theology, which reasons from nature to the existence and attributes of God. ID does not claim that all species of living things were created in their present forms, and it does not claim to provide a complete account of the history of the universe or of living things.
ID also is not considered by its theorists to be an “argument from ignorance”; that is, intelligent design is not to be inferred simply on the basis that the cause of something is unknown (any more than a person accused of willful intent can be convicted without evidence). According to various adherents, ID does not claim that design must be optimal; something may be intelligently designed even if it is flawed (as are many objects made by humans).
ID may be considered to consist only of the minimal assertion that it is possible to infer from empirical evidence that some features of the natural world are best explained by an intelligent agent. It conflicts with views claiming that there is no real design in the cosmos (e.g., materialistic philosophy) or in living things (e.g., Darwinian evolution) or that design, though real, is undetectable (e.g., some forms of theistic evolution). Because of such conflicts, ID has generated considerable controversy.